Plant cell:西南大学何光华/张婷团队发现控制水稻叶片宽度的分子机制
时间: 2022-07-31 点击次数:次 作者:
来源:Science Art
研究背景
叶片形态是理想植物结构的最重要特征之一。然而,控制作物这一特性的遗传和分子机制在很大程度上仍不清楚。由于叶子对光合作用很重要,叶片宽度是植物构建的关键组成部分。适当的叶宽在提高光能的吸收和转换效率方面具有重要的生物学意义(Ort等,2015)。因此,深入研究叶宽的调控机制不仅在理论上很重要,而且在提高水稻(Oryza sativa)产量方面也很重要。叶子从茎尖分生组织发育而来,茎尖分生组织在其外围含有专门的快速分裂细胞,首先发育成叶原基,然后改变它们的分裂平面并沿近端/远端,近轴/远轴和内侧建立三维轴/横轴(Fleming,2005;Moon和Hake,2011)。生长素通过两个影响生长素极性运输和合成的窄叶基因NAL1(窄叶1)和NAL7帮助调节叶宽。NAL1编码几乎未知生化功能的植物特异性蛋白质。NAL1的突变显着降低了极性生长素的转运能力,从而改变了维管组织的分布模式并减小了叶宽(Qi等,2008;Chen等,2012)。相比之下,NAL7编码含黄素的单加氧酶,其与丝兰属具有序列同源性并参与生长素生物合成。nal7突变体显示生长素含量降低和叶片表型狭窄(Fujino等,2008)。
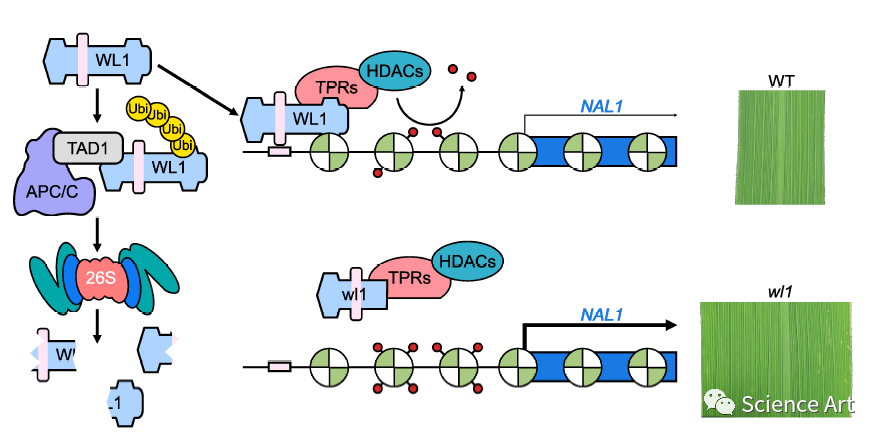
C2H2锌指蛋白家族是一类重要的真核转录因子,涉及涉及植物发育和植物应激反应的多种细胞过程(Sun等,2010)。DST(干旱和耐盐性)编码新的C2H2锌指转录因子,其在植物应激反应中极为重要。DST通过新的信号转导途径直接调节与H2O2稳态相关的基因,从而负调节气孔关闭。DST功能的丧失促进了气孔关闭并降低了气孔密度,导致水稻的干旱和耐盐性增强(Huang et al。,2009)。DST还通过与共激活因子DST共激活因子1(DCA1)相互作用来调节干旱和耐盐性,以调节编码H2O2清除剂的过氧化物酶24前体(PRX24)的表达(Cui等,2015)。在这项研究中,我们分离了一个宽叶1(wl1)突变体,叶宽增加。我们报告wl1表型是由于DST中的一个新的等位基因突变。尽管WL1/DST在水稻中的干旱和耐盐功能已有报道(Huang et al。,2009;Li et al。,2013;Cui et al。,2015),但编码蛋白如何控制叶片宽度尚未详细研究。我们的研究为阐明APC/CTAD1-WL1-NAL1途径介导的水稻叶片宽度控制建立了重要的遗传和分子框架,从而表明它对调节作物叶片结构至关重要。

研究结果
在这里,我们对水稻(Oryza sativa)宽叶1(wl1)突变体进行了表型分析,该突变体的叶片比野生型(WT)更宽,因为其维管束更多,小维管束之间的距离更大。WL1编码一种Cys-2/His-2型(C2H2)锌指蛋白,该蛋白与分蘖和矮秆1(TAD1)相互作用,后者是后期促进复合物/环体(APC/C)(一种多亚基E3连接酶)的共同激活剂。APC/CTAD1复合物通过泛素-26S蛋白酶体降解途径降解WL1。TAD1的功能丧失导致植物叶片狭窄,这是由于维管束数量减少和维管束之间的距离减小。有趣的是,我们发现WL1通过招募辅阻遏物TOPLESS-RELATED PROTEIN 并直接结合到NAL1调节区,减少染色质组蛋白乙酰化来抑制其表达,从而负调控窄叶基因窄叶1(NAL1)的表达。此外,生化和遗传分析表明,TAD1、WL1和NAL1通过共同的途径控制叶宽。我们的研究为理解APC/CTAD1-WL1-NAL1途径介导的水稻叶宽控制奠定了重要框架,并为改善作物结构提供了见解。

原文链接:https://academic.oup.com/plcell/advance-article/doi/10.1093/plcell/koac232/6651849?login=true

官方微信公众号
Copyright © 2011 Auto Parts All Right Reserved https://rice.sicau.edu.cn 版权所有© 四川农业大学水稻研究所
地址:四川省成都市温江区惠民路211号 邮编611130 联系电话:028-86290903、 028-86290897 邮箱:sdsb2006@163.com