水稻光敏雄性核不育作用机理
时间: 2016-12-15 点击次数:次 作者:admin
水稻(Oryza sativa L.)是世界上特别是中国最主要的粮食作物之一,养活了世界上近三分之一的人口。水稻雌雄同花,属于自花授粉作物,以自交结实为主。千百年来水稻的种植都是自交收种,依靠自然突变和人工选择获得对当地自然和栽培环境具有良好适宜性的优良水稻品种,以提高水稻单株产量。上世纪50年代的第一次绿色革命利用矮杆基因培育出了矮秆、耐肥、抗倒伏的高产水稻,使得世界粮食总产量有了大幅提升。这种产量的提升还是基于水稻的自交结实,产量的提高潜力有限,但某些水稻品种之间进行杂交能获得超过自交近20%的产量优势。水稻特殊的花结构导致去雄困难,不可能在生产上大规模制备杂交种。直到1966年袁隆平提出了通过选育不育系、保持系和恢复系的三系配套法来实现杂交稻的生产设想,并在10年后应用于生产中,开始在全国大面积种植杂交稻。从那时起杂交稻在我国的种植面积逐年提升,到上世纪90年代至今一直超过了总种植面积的50%。
三系配套法中的不育系属于细胞质雄性不育(Cytoplasmic male sterility,CMS),植物的雄性不育现象还存在细胞核雄性不育。细胞核雄性不育通常由隐性基因控制,在生产上难以找到保持系,保持其后代的不育性,因此难以利用。直到1973年石明松发现了光敏雄性不育株农垦58S,农垦58S具有长日照下不育、短日照下可育的特性,利用这一特性在长日照下可以作为不育系,在短日照下作为保持系,一系两用,大大简化了生产流程(BioArt注:石明松作为项目第二完成人(第一完成人为袁隆平院士)荣获2013年国家科学技术奖特等奖,然而此时他已经意外辞世26年了。1973年他发现的三株雄性不育系以后照亮了整个水稻世界,值得铭记)。后来我国育种学家们又找到了一类育性受温度影响的温敏雄性不育系,这些环境敏感性的细胞核不育系(environment-sensitive GMS,EGMS)是两系配套法利用的核心。相对于三系配套,两系不育系配套组更自由、恢复系广、杂交制种过程简化,在杂交稻生产上占据了越来越重要的地位。对于细胞质雄性不育的机理已经研究的较为清楚,但对于环境敏感型的细胞核不育系的作用机理的研究则相对来说比较少。
水稻(Oryza sativa L.)是世界上特别是中国最主要的粮食作物之一,养活了世界上近三分之一的人口。水稻雌雄同花,属于自花授粉作物,以自交结实为主。千百年来水稻的种植都是自交收种,依靠自然突变和人工选择获得对当地自然和栽培环境具有良好适宜性的优良水稻品种,以提高水稻单株产量。上世纪50年代的第一次绿色革命利用矮杆基因培育出了矮秆、耐肥、抗倒伏的高产水稻,使得世界粮食总产量有了大幅提升。这种产量的提升还是基于水稻的自交结实,产量的提高潜力有限,但某些水稻品种之间进行杂交能获得超过自交近20%的产量优势。水稻特殊的花结构导致去雄困难,不可能在生产上大规模制备杂交种。直到1966年袁隆平提出了通过选育不育系、保持系和恢复系的三系配套法来实现杂交稻的生产设想,并在10年后应用于生产中,开始在全国大面积种植杂交稻。从那时起杂交稻在我国的种植面积逐年提升,到上世纪90年代至今一直超过了总种植面积的50%。
三系配套法中的不育系属于细胞质雄性不育(Cytoplasmic male sterility,CMS),植物的雄性不育现象还存在细胞核雄性不育。细胞核雄性不育通常由隐性基因控制,在生产上难以找到保持系,保持其后代的不育性,因此难以利用。直到1973年石明松发现了光敏雄性不育株农垦58S,农垦58S具有长日照下不育、短日照下可育的特性,利用这一特性在长日照下可以作为不育系,在短日照下作为保持系,一系两用,大大简化了生产流程(BioArt注:石明松作为项目第二完成人(第一完成人为袁隆平院士)荣获2013年国家科学技术奖特等奖,然而此时他已经意外辞世26年了。1973年他发现的三株雄性不育系以后照亮了整个水稻世界,值得铭记)。后来我国育种学家们又找到了一类育性受温度影响的温敏雄性不育系,这些环境敏感性的细胞核不育系(environment-sensitive GMS,EGMS)是两系配套法利用的核心。相对于三系配套,两系不育系配套组更自由、恢复系广、杂交制种过程简化,在杂交稻生产上占据了越来越重要的地位。对于细胞质雄性不育的机理已经研究的较为清楚,但对于环境敏感型的细胞核不育系的作用机理的研究则相对来说比较少。
近日,华中农业大学张启发课题组在2016年12月12日的PNAS上以“PMS1T,producing phased small interfering RNAs,regulates photoperiod-sensitive male sterility in rice”为题,报道了在水稻光敏型雄性不育作用机理中发挥作用的另一个基因PMS1T,该基因能够被miR2118识别并介导剪切,并从剪切位点开始形成一串21nt的phasiRNA,进一步研究发现这些phasiRNA在长日照下的农垦58S中的表达量明显高于正常可育的对照品种。不包含完整miR2118识别位点的PMS1T则不能发挥正常的功能来降低长日照下的育性,由此表明PMS1T经过miR2118介导产生的phasiRNA在调控水稻光敏雄性不育过程中发挥着重要作用。值得注意的是,PMS1T是到目前为止鉴定到的第一个具有生物学功能的PHAS基因(能产生phasiRNA的基因),证明了这类小RNA对植物生长发育的重要性。
其实在这篇文章背后,除了PMS1T基因功能及其发挥作用方式的研究亮点外,更值得关注的是对这个基因定位的艰辛。这项研究被张启发老师称之为一个“化石”类的研究——历史悠久,张老师课题组从1987年开始了光敏不育的相关研究,1994年定位到了这个基因位点pms1 (photoperiod-sensitive male sterility1,这是最开始的命名,之前报道pms3也是光敏不育的一个非常重要的基因,下文将提及),这期间一直试图克隆该基因。早在2001年就发表了对pms1的精细定位结果。通过籼型不育组合32001S/明恢63将pms1定位于两个分子标记Fssr和Rssr之间85 kb范围内。后来构建了覆盖该区间的6个互补载体转化农垦58S后,发现这6个片段均不能恢复长日照下农垦58S的育性(该结果未发表)。于是改用农垦58S/明恢63组合对pms1重新进行定位,最终将pms1定位在85 kb相邻的21 kb区间内,分段互补仍然没有能够得到预期的结果(该定位结果最终并未发表)。在这样的研究背景下,该篇论文的一作范优荣不得不考虑问题到底出在哪里,仔细分析了前人的研究结果后,认为可能是群体中复杂的遗传背景导致了定位的偏差。为了清除背景的影响,在农垦58S/明恢63组合中用农垦58S进行连续回交,最终通过对几个高世代回交F2群体的分析发现农垦58S中的pms1不是隐性,而是显性,并且这种光敏雄性不育特性表现为不完全显性。这也解释了之前无法通过转基因验证定位的结果:因为之前都是将隐性的pms1转化至显性的农垦58S中,那必然不会有效果。找对了方向,接下来精细定位的工作就进行得比较顺利了,经过两年多的努力,最终将pms1定位到一个3.8 kb的区间。将农垦58S的这段序列互补转化到近等基因系中能显着降低受体的育性,反过来将来自于明恢63的这段序列转化到农垦58S中对育性没有影响,说明定位区间内确实含有pms1的候选基因但在这区间内并没有发现任何预测的基因。通过prime walking以及5’RACE和3’RACE的方法,最终得到了pms1的全长cDNA,仅包含一个exon,随后将该转录本命名为PMS1T。对PMS1T进行RNAi和超量表达证实了就是pms1基因。PMS1T及其启动子区在58S和明恢63之间仅存在四种序列差异,随后通过遗传分析、比较测序和转基因验证排除了其中三个变异位点,最终确定一个SNP的变异(不育为“T”,可育为“G”)导致了功能的差异。
其实研究做到这里才只进行了一半,而这一半却耗费了几届硕博研究生生的努力,三次精细定位,走了很多弯路。后续的研究,面对这样一个完全未知的基因,如何解析它的功能又是一项挑战。2012年张老师课题组对于另一个光敏不育相关基因pms3的定位克隆发现:由于一个SNP的差异(农垦58为“G”,农垦58S为“C”)引起pms3编码的1,236 bp长的long non-coding RNA,LDMAR的RNA二级结构发生了改变,造成其启动子区域DNA甲基化程度的升高,抑制了基因在长日照下幼穗中的表达量,导致雄性不育。进一步研究发现LDMAR启动子产生的21-nt的小RNA Psi-LDMAR介导了该区间的DNA甲基化。后来华南农业大学庄楚雄和刘耀光课题组的研究结果与此不谋而合,不过他们认为SNP附近产生的一个21 nt的小RNA(SNP位于小RNA的第11位上)可能更重要。两个不同的课题组运用不同的光温敏材料定位到了同一个基因,也同时解析了相似的作用机制。还是蛮有意思的。
回到开头,在这项研究中作者起初也认为可能pms1的作用机制与pms3类似,因为实验表明pms1也编码一个lncRNA。但后续的实验证明,两者作用方式并不相同。序列分析发现PMS1T的5’端存在miR2118的识别位点,接下来的实验也证实了miR2118能够结合到PMS1T上,并介导对PMS1T的剪切。而水稻中miR2118主要参与介导形成21-nt的phasiRNA,miR2118结合到靶基因上后,会介导靶基因从剪切位点开始形成一串以21-nt为相位连续排列的小RNA,这类小RNA就称之为phasiRNA。
小RNA测序的数据发现在PMS1T上从miR2118剪切位点开始确实能形成一系列的phasiRNA,共计18对。这些phasiRNA在体内是真实存在的,并且在长日照的农垦58S中表达量明显高于对照材料,表明phasiRNA与光敏雄性不育相关,分析转基因材料中phasiRNA的表达量进一步证实了该结论。而此前鉴定到的功能性碱基突变位点就位于miR2118剪切位点后24 bp上可能导致在长日照的农垦58S中能产生更多的phasiRNA,一方面这些累积的phasiRNA可能造成了雄性不育;另一方面phasiRNA可能影响下游一些未知靶基因的表达从而导致了光敏雄性不育。
故事很长,也很精彩,十年的努力没有白费,最终向人们揭示了一个物种进化过程又一个神奇的瞬间。但故事并没有结束,因为我们并不知道这些产生的phasiRNA到底如何发挥作用?
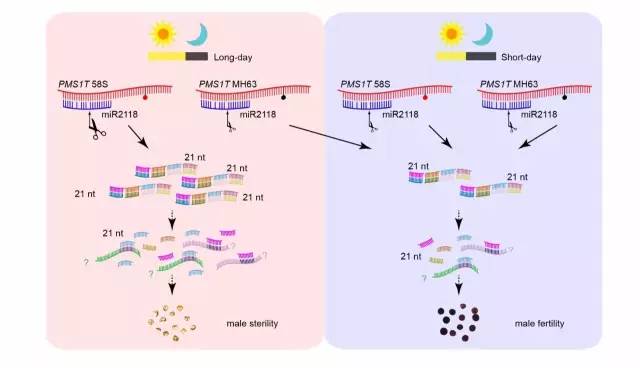
pms1 调控光敏感雄性核不育的作用机理
来源:网络,撰文丨宗伟 (华中农业大学生命科学学院博士)

官方微信公众号
Copyright © 2011 Auto Parts All Right Reserved https://rice.sicau.edu.cn 版权所有© 四川农业大学水稻研究所
地址:四川省成都市温江区惠民路211号 邮编611130 联系电话:028-86290903、 028-86290897 邮箱:sdsb2006@163.com